
Formering, forplantning, dannelse af nye individer, afkom, af allerede eksisterende individer; et fællestræk for alt levende og den måde, hvorpå arter eksisterer i tidsrum længere end det enkelte individs livslængde. Siden Louis Pasteur i 1860'erne definitivt tilbageviste idéen om spontan genese, dvs. at levende organismer skulle kunne opstå spontant af ikke-levende materiale eller af levende materiale af anden art, har det været en basal biologisk antagelse, at levende organismer kun frembringes af andre levende organismer: omne vivum ex vivo 'alt levende stammer fra det levende'.
formering (forplantning)

Formering. Bakterier formerer sig altid ukønnet, men det kan ske på flere måder. Bakterier kan også udveksle arvemateriale ved konjugation.

Formering. Eksempler på kønnet og ukønnet formering hos dyr med eller uden generationsskifte.
Ukønnet og kønnet formering
Ukønnet formering er den evolutionshistorisk ældste formeringsmåde. I sin simpleste form består den blot i opsplitningen af et individ i nye, selvstændige individer; som fx en bakteriecelles deling. Men der er hos mange arter udviklet yderst avancerede træk og adfærdsmønstre i forbindelse med ukønnet formering, fx særlige vegetative spredningsenheder hos planter og knopskydning hos søanemoner.
Ukønnet formering er en yderst effektiv formeringsform, der gør det muligt for enkeltindivider at producere store mængder afkom på relativt kort tid; ud fra et resursemæssigt synspunkt er ukønnet formering den kønnede langt overlegen. Ukønnet formering kendes fra næsten alle organismegrupper, fx findes der krybdyr og fisk, som formerer sig ukønnet ved partenogenese, jomfrufødsler, vha. ubefrugtede æg; kun blandt pattedyr og fugle er ukønnet formering ukendt.
Kønnet formering
Den kønnede formering omfatter to vigtige elementer, nemlig en befrugtning og en reduktionsdeling, meiose, der giver mulighed for nye kombinationer af gener fra forældrenes arvemasser (genomer), en såkaldt rekombination. Befrugtningen består oftest af en fusion af to kønsceller, gameter, efterfulgt af en fusion af de to haploide kerner i kønscellerne. Herved opstår en zygote, hvis diploide kerne indeholder begge kønscellernes kromosomsæt. På et eller andet tidspunkt i de kønnede organismers livshistorie sker der en reduktionsdeling, hvorved cellerne igen bliver haploide. Kønnet formering er således forbundet med et kernefaseskifte. Ved ukønnet formering har formeringsenhederne derimod samme kernefase som moderindividet (haploid eller diploid), fordi de dannes ved mitotiske kernedelinger. Se også celledeling og generationsskifte.
Årsager til den kønnede formerings udbredelse
Det kan undre, at kønnet formering med en 50:50 kønsfordeling er så almindelig en forplantningsmåde især blandt dyr, da den rent numerisk er mindre effektiv end den ukønnede, idet oftest kun hunnens resurser bruges på produktion af afkom, hvis halve genom oven i købet stammer fra et andet individ. En mulig forklaring er, at den opblanding af arveanlæg fra forskellige individer, der sker ved kønnet formering, resulterer i stor genetisk variation og dermed en evolutionær fordel gennem bedre muligheder for tilpasning til ændringer i miljøet.
Konjugation
En særlig formeringsmåde er konjugation hos ciliater. Her sker en delvis fusion mellem to celler af forskellig parringstype, som derefter gensidigt udveksler haploide kerner, der smelter sammen med en stationær kerne, hvorefter de to individer adskilles.
Blandt prokaryoter, bl.a. bakterier, er konjugation betegnelsen for udveksling af genetisk materiale mellem fusionerende celler. Dette fænomen er baggrund for overførslen af antibiotikaresistens mellem forskellige bakterier og repræsenterer derved et alvorligt sundhedsproblem.
Formering hos dyr
Dyr kan danne afkom ved kønnet eller ukønnet formering. Ved ukønnet formering får afkommet samme arvemasse som moderindividet. Ved kønnet formering blandes to forældreindividers arvemasse ved befrugtning.
Ukønnet formering
Ukønnet formering ved deling af modercellen i to identiske datterceller er den dominerende formeringsmåde hos encellede organismer. Der findes dog også tilfælde, hvor modercellen vokser sig stor og derefter deler sig i et stort antal datterceller; det er fx tilfældet for malariaparasitten, en sporozo, hvilket fremkalder de rytmiske feberanfald hos malariapatienter.
Hos flercellede dyr er det cellekomplekser eller fuldt færdige individer, der frigør sig fra moderindividet ved ukønnet formering, hvilket både kan ske i fosterstadiet, i forskellige ungdomsstadier samt hos voksne. Enæggede tvillinger, der hos mennesket er et sjældent fænomen, er et eksempel på ukønnet formering i fosterstadiet; hos det niringede bæltedyr er det normen, at zygoten deler sig to eller tre gange og giver ophav til fire eller otte genetisk ens unger. Lignende former for polyembryoni kendes bl.a. fra visse snyltehvepse. Ukønnet formering kan også forekomme i livsstadier forskellige fra det kønsmodne; det gælder fx for de parasitiske fladorme ikter og bændelorme, visse cølenterater samt hos salper (en chordatgruppe, se sækdyr).
Nye individer kan dannes på mangfoldige måder ved både indre og ydre knopskydning, og ukønnet formering er i det evt. kønsmodne voksenstadium ofte forbundet med stor regenerationsevne. Knopskydning kan være svær at skelne fra deling. Visse fladorme og børsteorme deler sig uden forberedelse, og de manglende organer gendannes bagefter, architomi. Andre orme og fx søanemoner nydanner organer, før individerne adskilles, såkaldt paratomi. Hos nogle cølenterater og sækdyr samt mosdyr adskilles individerne ikke, men forbliver samlede i kolonier med mere eller mindre intim forbindelse mellem enkeltdyrene og ofte med en vis arbejdsdeling mht. fødeoptagelse, forsvar mv.
Ferskvandssvampe og ferskvandsmosdyr kan danne særlige kimknopper, gemmulae hhv. statoblaster. I disse er levende kimceller omgivet af en beskyttende skal, der muliggør overlevelse under ugunstige kår og fremmer spredningen.
Ukønnet formering kan også ske ved partenogenese. Dette kendes fx fra mange insekter, fisk og krybdyr, hvor alle eller de fleste individer er hunner. I nogle tilfælde kan arter veksle mellem kønnet formering og partenogenese. Visse partenogenetiske sydamerikanske tandkarper parrer sig med hanner af nærtstående arter, da ægget kun kan påbegynde sin udvikling efter påvirkning fra sædceller.
Kønnet formering
Ved dyrs befrugtning forenes arvemassen i en hunlig ægcelle, oocyt, som er dannet i en kønsmoden æggestok, ovarium, med arvemassen i en hanlig sædcelle, spermatozo, som er dannet i en kønsmoden testikel, testis. Læs mere om den kønnede formering hos dyr og menneske.
Sporeplanter
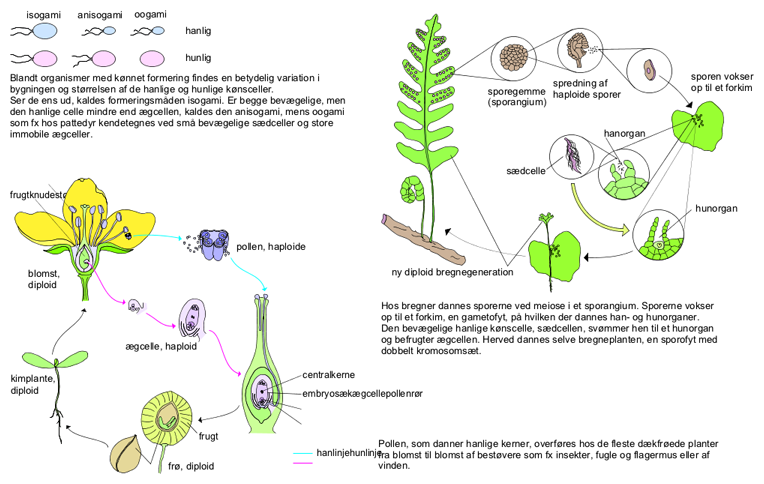
Formering. Livscyklussen hos mange frøplanter og sporeplanter som fx bregner omfatter både en kønnet og en ukønnet generation.
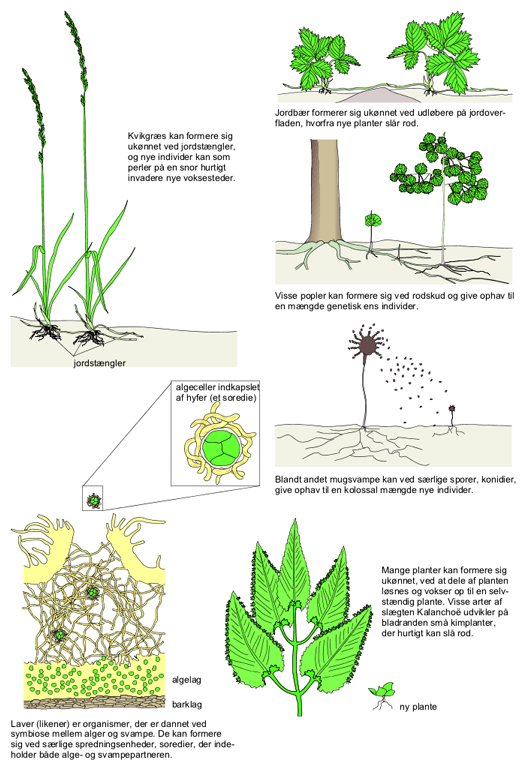
Formering. Mange planter, svampe og laver kan formere sig ukønnet og det på et utal af måder. Haveplanter og fx frugttræer formeres ofte ukønnet; i princippet er deling af en staude til udplantning blot endnu et eksempel herpå, blot med menneskers hjælp.
Hos sporeplanter findes både ukønnet og kønnet formering, der begge har til formål at danne nye individer. Disse udvikles ofte fra særlige formeringsenheder, diasporer, som også tjener til spredning af den enkelte art.
Ukønnet formering
Den simpleste form for ukønnet formering, mitotisk celledeling, hvorved en modercelle deles i to datterceller, er den hyppigste hos encellede alger og svampe. Mange flercellede alger og laver formerer sig ved fragmentering, hvor dele af individet rives løs og siden vokser ud til nye individer. I nogle tilfælde er fragmenterne særligt udformede organer: Mange laver danner isidier og soredier, som består af både alge- og svampepartneren i laven, og en del mosser og bregner danner knopkorn og ynglelegemer.
Det særlige kendetegn ved sporeplanter er deres evne til at danne sporer, som kan være kønnede eller ukønnede. De ukønnede sporer er en- eller fåcellede. Nogle dannes i særlige beholdere, sporangier; disse sporer er enten ubevægelige aplanosporer eller bevægelige zoosporer, som er forsynet med svingtråde. Begge typer er nøgne, uden cellevæg. Zoosporer er almindelige hos ægsporesvampe samt mange brun- og grønalger. Mange svampe, især sæksporesvampe, formerer sig ved konidier, som er vægklædte sporer, der dannes uden på hyferne, ofte i stort antal. Endelig findes en del tykvæggede hvilesporer, som er specielt egnede til at overleve ugunstige perioder og desuden fungerer som spredningsenheder, fx klamydosporer hos svampe samt akineter og cyster hos alger. Hos nogle sporeplanter, fx øjealger samt mange mug- og skimmelsvampe, kender man kun ukønnet formering.
Kønnet formering
Første led i en kønnet formering, befrugtningen, består grundlæggende af en fusion af to haploide kerner; denne fusion kan hos sporeplanter tilvejebringes på adskillige måder. Den mest udbredte befrugtningsmåde er fusion af to kønsceller, gameter. Gameterne kan være bevægelige eller ubevægelige og dannes oftest i særlige kønsorganer, gametangier. Hos encellede flagellater kan den bevægelige celle dog selv fungere som gamet.
Man skelner mellem tre typer af gametbefrugtning, gametogami: isogami, anisogami og oogami. Ved isogami er gameterne ens af udseende og størrelse, og de kan derfor ikke henføres til et bestemt køn. Hos nogle arter kan gameter fra samme individ smelte sammen, men oftest stammer de fra genetisk forskellige individer, såkaldte parringstyper, der ofte betegnes + og −. Ved anisogami har gameterne samme udseende, men er ikke lige store; den mindste og ofte den mest aktive er da en hanlig gamet, den store en hunlig. Den mest avancerede type er ægbefrugtningen, oogami, hvor fusionen sker mellem en stor, ubevægelig ægcelle og en væsentlig mindre, hanlig gamet, som kan være bevægelig, et spermatozoid, eller ubevægelig, et spermatium. Det hanlige kønsorgan kaldes oftest et antheridium, det hunlige et oogonium. Hos mosser og karsporeplanter ligger ægcellen inde i et særligt, flaskeformet organ, et arkegonium.
Hos alger er gametbefrugtning enerådende, og alle de nævnte typer findes repræsenteret, også inden for enkelte klasser som fx brunalger og grønalger; hos svampe er dette kun tilfældet for Chytridiomycetes. Oogami er enerådende hos rødalger, mosser og karsporeplanter. De anisogame og oogame arter kan være enbo, dvs. kønsorganerne findes på samme individ, eller tvebo, dvs. der findes adskilte han- og hunplanter. Begge typer forekommer hos alger og mosser, mens de fleste karsporeplanter er enbo.
En befrugtning, dvs. en kernefusion, kan imidlertid godt finde sted, uden at der dannes kønsceller. I stedet kan der ske en fusion af flerkernede gametangier, gametangiogami, som det er tilfældet hos mange svampe. Hos koblingssvampe er gametangierne ens, og kernerne fra hvert gametangium samles parvis i den opståede zygote, zygosporen, og formeringen kaldes da zygogami. Kernefusionen finder sted enten i zygoten eller på et senere tidspunkt. Hos sæksporesvampene dannes der to forskellige gametangier, et antheridium og et ascogonium; efter fusionen af gametangierne placerer kernerne sig parvis i ascogoniet, og kernefusionen indtræffer først senere. Hos ægsporesvampe vandrer kerner fra et antheridium over i et oogonium, hvor kernerne straks fusionerer parvis og danner flere zygoter, oosporer.
De fleste svampe med gametangiogami er enbo, men blandt disse skelner man mellem to typer af forligelighed, kompatibilitet. Nogle svampearter er homothalliske, dvs. kønnet formering kan ske mellem alle mycelier eller inden for samme mycelium; andre er heterothalliske, dvs. mycelierne skal være af forskellig parringstype.
Befrugtningen kan endvidere ske ved fusion af uspecialiserede, vegetative celler. Denne form for befrugtning, somatogami, spiller en betydelig rolle hos svampene og er typisk for stilksporesvampene, hvor fusionen sker mellem to forligelige mycelier. Også her er der to typer af forligelighed, men 90 % af alle stilksporesvampe er heterothalliske.
Livshistorier
En kønnet formering indebærer som tidligere nævnt et skifte mellem det haploide og det diploide kromosomtal, et kernefaseskifte, fordi der på et eller andet tidspunkt efter befrugtningen sker en reduktionsdeling, meiose, af den diploide kerne i zygoten. En organismes livshistorie afhænger af, hvornår denne reduktionsdeling finder sted. På basis heraf kan man opstille tre hovedtyper, som sammen med diverse varianter alle er repræsenteret hos sporeplanterne.
Hos haplonter sker reduktionsdelingen ved spiring af zygoten. Individerne er haploide, og zygoten er den eneste diploide celle. Denne type findes fx hos de fleste grønalger og hos nogle koblingssvampe. Hos diplonter sker reduktionsdelingen ved dannelse af gameter. Individerne er diploide, og kun gameterne er haploide. Denne type forekommer langt mere sporadisk i planteriget end i dyreriget, hvor den er næsten enerådende, men alle ægsporesvampe og kiselalger samt brunalgen klørtang er diplonter.
Den tredje type, haplodiplonterne, er kendetegnet ved, at der sker et generationsskifte mellem to typer af planter, en haploid gametofyt og en diploid sporofyt. Gametofytten danner haploide gameter, der fusionerer til en diploid zygote; denne spirer op til en sporofyt, som ved meiose danner haploide sporer, ofte betegnet meiosporer. Fra disse meiosporer dannes nye gametofytter, og livshistorien er sluttet.
Hermed er variationen dog ikke slut. Der findes nemlig to undertyper af haplodiplonter, afhængigt af hvorledes gametofytten og sporofytten er bygget. Hvis de er ens, kaldes generationsskiftet isomorft, er de forskellige heteromorft. Et heteromorft skifte findes hos alle nulevende mosser og karsporeplanter, men hvilken generation, der udgør den store, grønne plante, varierer; hos mosserne er det gametofytten, hos karsporeplanterne (fx bregner) er det sporofytten. Blandt algerne kendes både isomorft og heteromorft generationsskifte hos mange brunalger og visse grønalger; de fleste rødalger er også haplodiplonter, men deres livshistorie er speciel ved, at der findes to forskellige sporofytter, en snyltende og en fritlevende. Hos svampe er generationsskifte sjældent, men findes fx hos chytridiomyceten Allomyces.
Endelig findes to typer af livshistorier, som kun forekommer hos svampe. De er karakteriseret ved, at der mellem den haploide og den diploide fase er indskudt en parkernefase, dikaryofase, hvor to haploide kerner ligger ved siden af hinanden og deler sig samtidig. Kernefusionen sker først senere, umiddelbart inden reduktionsdelingen og dannelsen af svampens meiosporer, dvs. kun zygotekernen er diploid. Hos sæksporesvampe dominerer det haploide mycelium, mens dikaryofasen er indskrænket til en lille del af frugtlegemet; hos stilksporesvampe er dikaryofasen den mest dominerende, idet størstedelen af myceliet og hele frugtlegemet er dikaryotisk.
Frøplanter
Også hos frøplanter findes såvel ukønnet som kønnet formering. Mange af de planter, vi benytter i have- og landbrug, formeres ved frøformering. Læs mere om formering hos frøplanter.
Kommentarer (1)
skrev Lars Jødal
Ifølge Lex-opslag for "cølenterater" er denne betegnelse forældet. Skal disse sætninger måske formuleres anderledes?
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.