| Boks 7-5: Smådyr i skovens luftrum
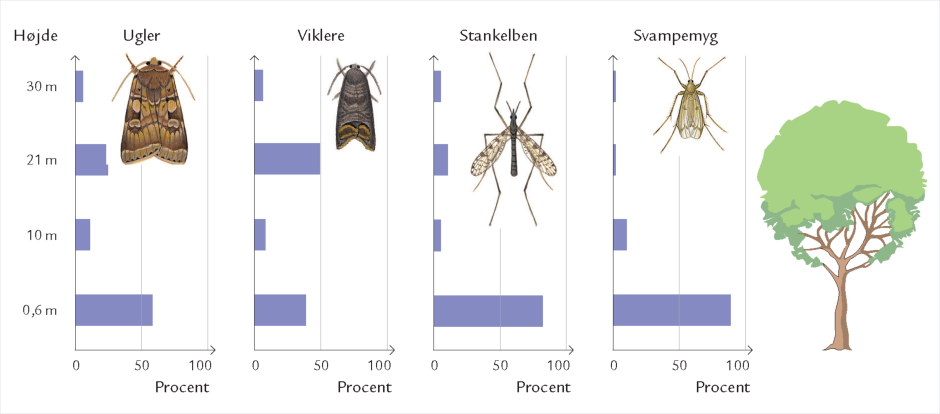
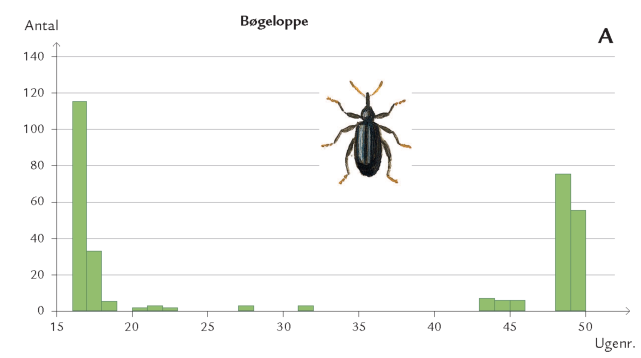
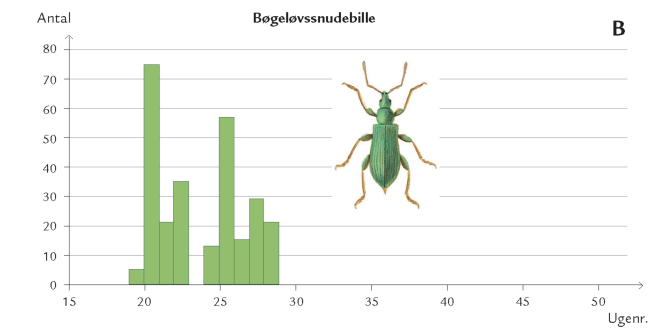
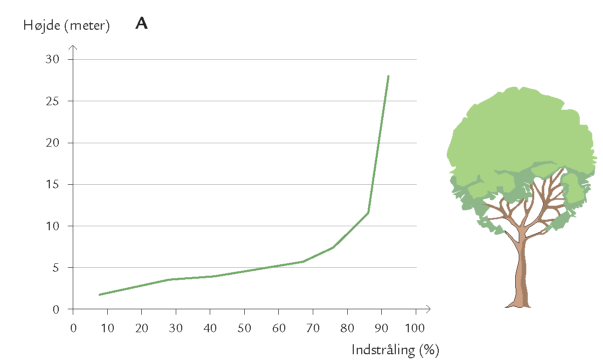
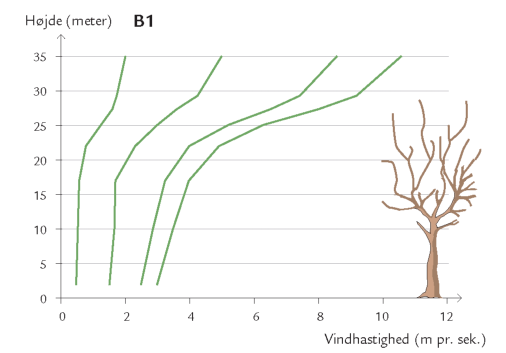
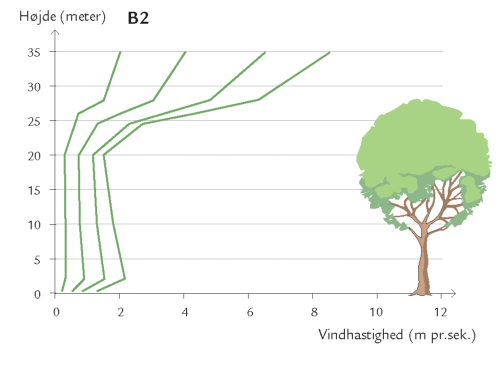
Døgnet rundt er insekter på vingerne i luftrummet mellem skovens stammer og trækroner – på jagt efter føde og ynglesteder eller som led i parrings- eller spredningsadfærd. Undersøgelser i Hestehaven har vist, at det er forskelligt, hvor højt over skovbunden de forskellige insektarter flyver. Højden afhænger af insekternes størrelse og flyveevne i relation til vindpåvirkningen i de forskellige højder (figur 7-10) samt af arternes biologiske tilknytning til bestemte lag i skoven. Hvor yngler de? Hvor finder de værtsplanten? Hvad lever de af? Generelt aftager svagt flyvende skovbundsinsekters flyveaktivitet med højden. Det gælder f.eks. små mygge- og billearter (figuren nedenfor).
Vindspredning af smådyr Også smådyr uden vinger kan krydse skovens luftrum, f.eks. kan nyklækkede frostmålerlarver – se længere nede i artiklen – ved vindens hjælp spredes fra træ til træ eller til andre bevoksninger, selv ved meget svage vinde.
Heller ikke edderkopper kan flyve ved egen kraft, men ved at bruge en silketråd til at bære sig gennem luften, kan de spredes til nye levesteder. Edderkopper er ofte de første dyr, der koloniserer nye landområder i kraft af denne effektive spredningsteknik. Der er mange forskellige teknikker, men grundlæggende foregår det på den måde, at edderkoppen bevæger sig til et forholdsvis højtliggende punkt, strækker benene, retter bagkroppen opad og lader en eller flere silketråde flyde ud af spindevorterne. Silken løftes af opadstigende luftstrømme og kan nu – som en luftballon – bære edderkoppen til nye jagtmarker. Dette fænomen kaldes flyvende sommer, selv om det finder sted på næsten alle tider af året.
Hos større arter af edderkopper er det kun de unge stadier, der kan spredes på denne måde. De største massespredninger foretages af de små tæppespindere (familien Linyphiidae), der er meget almindelige i skove. På dage med opadstigende, varme luftstrømme kan de spredes i massevis og næsten fuldstændigt dække et område med et tæppe af silke.
|
Afsnit fortsætter her.
Barkens epifytter
Epifytter er organismer, der er specialiseret til at vokse i det meget udsatte miljø, barken på et levende træ udgør, og findes ikke mindst blandt mosser og laver. Det er især store svingninger i fugtigheden, der stiller store krav til epifytternes evne til at modstå udtørring. Skorpeformede laver udviser den stærkeste tilpasning til disse forhold og kan findes under ganske tørre forhold, mens buskformede laver og mosser generelt stiller lidt større krav om en rimelig fugtighed og derfor især findes i skove med et fugtigt mikroklima.
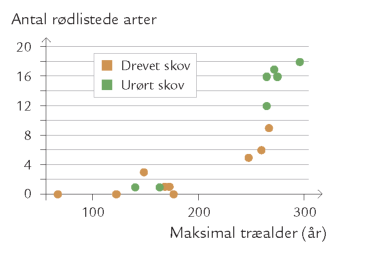
Både gamle og unge træer kan være voksested for epifytter, men de rigeste samfund findes i gamle skove (figur 7-33, se også afsnittet Veterantræer og dødt ved (inkl. følgende afsnit)). Det skyldes bl.a., at gamle træer har den groveste bark. Det giver bedre muligheder for tilhæftning, og den grove og tykke bark med uregelmæssige, dybe sprækker er langt bedre til at holde på fugtigheden end den glatte bark på yngre træer. Desuden er epifytsamfundene ikke isolerede fra livet i træets indre.
Hulheder fyldt med formuldede ved- og insektrester kan give anledning til en lokal næringsberigelse af det regnvand, der siver ned ad stammen og gennem hulheder i træets indre. På bøg findes adskillige sjældne arter således helt karakteristisk nedstrøms for små, ofte næppe synlige hulheder, der står i forbindelse med større hulheder inde i træet, hvor vedboende svampe sammen med insekter forårsager dannelsen af et næringsrigt, formuldet træsmuld (figur 7-34).
Smådyr på barkoverfladen
For de mange insekter, der skifter “lag” i løbet af deres udvikling, er de levende stammer en vigtig forbindelsesvej mellem skovbund og trækroner med et årstidsbestemt trafikmønster. I Hestehaven blev denne trafik overvåget ved hjælp af særlige fælder på bøgestammerne. Især om foråret går trafikken overvejende fra skovbunden mod trækronerne og består af insekter på vej fra udklækningsstederne i jordbunden til kronelaget, hvor de æder blade eller lægger æg (figur 7-35). I andre perioder vender trafikken, og de fleste insekter søger nu mod jordbunden, nogle for at lægge æg, andre for at forpuppe sig eller overvintre. Tilsvarende årstidsbestemte vandringer er observeret på stammer af ask og eg.
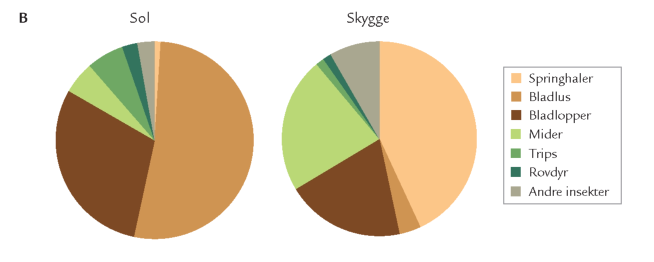
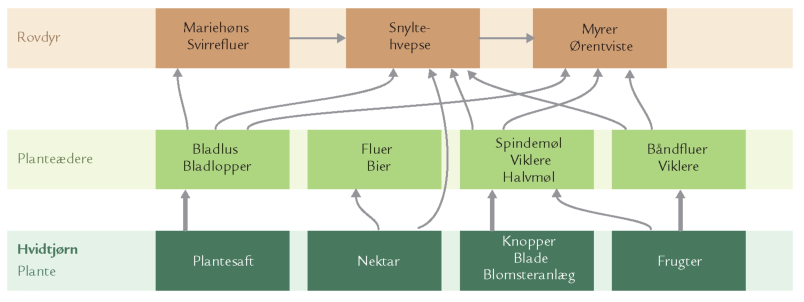
Epifytter spiller ingen særlig rolle for træernes tilvækst og økosystemets funktion i danske skove, men de er vigtige for dyrelivet i stammelaget. Bevoksninger af pulveralger, mos og lav på stammer af f.eks. bøg, eg og ask er således udgangspunkt for græsningsfødekæder (figur 7-1). I mos- og lavbevoksninger findes mængder af mider og springhaler, og på stammerne piler små barklus rundt (figur 7-36). I sensommernætter kan man se skovørentvisten (Chelidurella acanthopygia) græsse på stammerne (figur 7-37).
Smådyr på barken efterstræbes af adskillige rovdyr, bl.a. edderkopper som barkjæger (Drapetisca socialis) (figur 7-38) samt stor springedderkop (Marpissa muscosa) (figur 7-39). Om natten kan stammerne afpatruljeres af løbebiller fra skovbunden, f.eks. Carabus-arter (se figur 7-23).
Anvendelse af indsamlingsudstyr, der er særlig effektivt til fangst af helt små dyr, har vist, at hovedparten af barkfaunaen på bøg og eg består af springhaler, mider, trips, barklus m.fl. I en ung egebevoksning (55 år) var op mod 90 % af de indsamlede dyr under 3 mm lange; springhaler dominerede klart. I fugtigt vejr udnyttes stammernes epifytter af endnu flere dyr, der vandrer op fra jordbund og urtelag, f.eks. snegle. I naturskov med mos- og lavbevoksede stammer findes mange mos- og lavædende mider, der i flere tilfælde – ligesom laverne selv – er gode gammelskovsindikatorer (se kapitlet Særlige levesteder i skovene).
Selve barken udnyttes kun i meget ringe grad som fødeemne for insekter, men den kan være gemmested for mange smådyr om vinteren; det gælder især den dybt furede egebark. De overvintrende smådyr i barkens revner er et velbesøgt spisekammer for bl.a. mejser og spætmejser.
Vedædere og saftsugere
Stammelagets enorme vedmasse udnyttes kun i meget ringe grad af insekter. Vedædere som f.eks. barkbiller er primært knyttet til svækkede, døende eller døde stammer (kapitlerne Dynamik og forstyrrelser i skoven og Særlige levesteder i skovene). Generelt kræver det kraftige, bidende munddele at binde an med det hårde ved, der samtidig er næringsfattigt og til og med vanskeligt at fordøje. Derfor har nogle insekter også et samarbejde med vednedbrydende svampe, som det er beskrevet nærmere i Nedbrydningens tidlige faser.
Nogle insekter borer deres veludviklede, sugende munddele ind gennem stammebarken og tapper den næringsrige saft fra sikarrene (kapitlet Skovens vækst). Et eksempel er bøgeskjoldlusen (Cryptococcus fagisuga). Den 0,4-0,8 mm lange, vingeløse hun sidder på bøgebarken med sin tynde snabel permanent indboret (figur 7-40). Bøgeskjoldlusen er et let bytte for rovinsekter som mariehøns og små, rovlevende galmyglarver, Lestodiplosis. Ved masseangreb af bøgeskjoldlus kan barken skades og sprække, efterfølgende kan træet angribes af vedborende biller, f.eks. gul vedborer (kapitlerne Dynamik og forstyrrelser i skoven og Særlige levesteder i skovene) og eventuelt bukke under.
At få adgang til det saftførende væv under barken kræver normalt, at insekterne har lange, sugende munddele, men under visse omstændigheder kan også mindre veludrustede insekter fouragere på træernes saft. Beskadiges barken, så safttransporten afbrydes, kan der opstå saftflåd, hvor næringsrig saft siver ud og løber ned ad barken. På grund af gæringsprocesser får saften en karakteristisk lugt, der bl.a. tiltrækker bestemte flue- og billearter. Ved saftflåd på f.eks. elm, eg og bøg optræder bl.a. larver af visse svirrefluearter, f.eks. træsaftsvirrefluer (Brachyopa), og voksne individer af elmesaftbillen (Nosodendron fasciculare) og glansbillearter.
Kronelagets organismer
Kronelagets mange fødeemner, blade, kviste, frugter og frø, er et meget vigtigt spisekammer for planteædende smådyr, der igen er en betydelig føderessource for rovdyr.
Bladene brødføder langt de fleste planteædere, og tilsyneladende er trækronernes enorme bladmasse en lettilgængelig og næsten ubegrænset føderessource. Bladkost er imidlertid vanskelig at udnytte effektivt, bl.a. på grund af de svært nedbrydelige sukkerstoffer (cellulose og hemicellulose), cellevæggene består af. Dem kan kun de færreste insekter fordøje. En større sommerfuglelarve, der gnaver af bladpladen, kan således kun udnytte indholdet i de bladceller, der tilfældigvis bliver flået op, for larvens tarmkanal mangler enzymer eller mikroorganismer, som kan nedbryde sukkerstofferne i de intakte cellevægge. Ydermere indeholder løvtræernes blade garvestoffer (tanniner), der smager ilde, hæmmer insekternes udnyttelse af fødens proteinindhold og binder sig til visse enzymer i planteædernes celler. Bladenes garvestoffer fungerer dermed som et kemisk forsvar mod insektangreb.
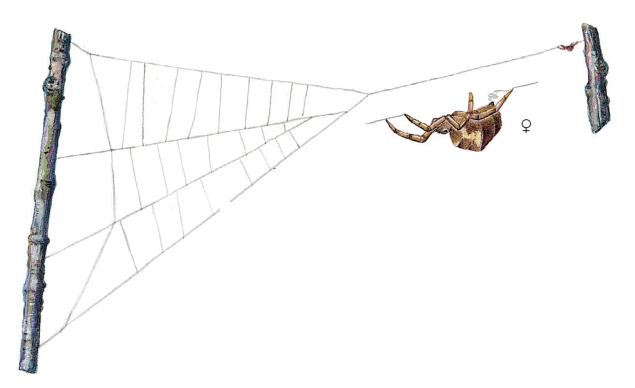
Kronelagets planteædere er som nævnt fødekilde for mange rovdyr, f.eks. bladlusædere som mariehøns, guldøjelarver og svirrefluelarver, men også for mange arter af edderkopper, f.eks. trekantspinderen (Hyptiotes paradoxus), der har et ret besynderligt udseende og en speciel jagtteknik. Den findes især i nåleskove, hvor den ofte bygger sit trekantede spind ret højt oppe mellem træernes grene. Edderkoppen sidder i den spidse vinkel af trekanten og holder fast i en silketråd, der bærer spindet i den ene side. Når et bytte går i nettet, slækker edderkoppen på spindet – og strammer det straks igen. På den måde får edderkoppen viklet sit bytte grundigt ind i nettet og kan nu mere sikkert nærme sig og fortære det. Trekantspinderen hører til en familie af edderkopper, Uloboridae, der er helt specielle ved at mangle giftkirtler. De må alene klare sig ved at pakke byttet grundigt ind i silke. Denne silke er svagt blålig og fungerer ligesom velcrotape. Deres spind er specielt ved ikke at være klistret ligesom f.eks. korsedderkoppens spind (figur 7-41).
Synkronisering med løvspring er livsvigtigt
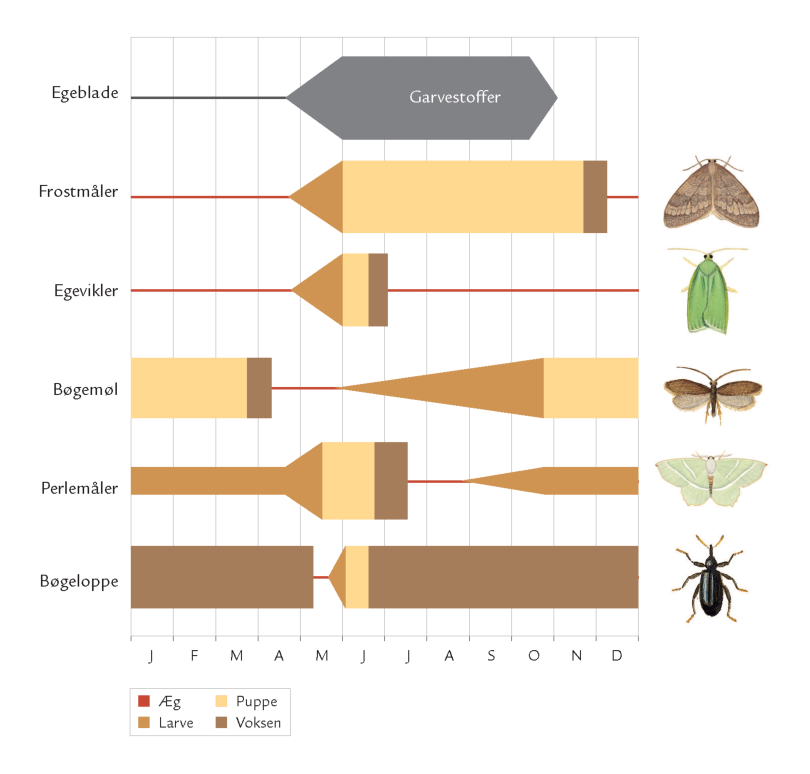
Efterhånden som løvtræernes blade ældes, bliver bladvævet endnu mere sejt og vanskeligt at udnytte, og et stigende indhold af garvestoffer nedsætter ligeledes fordøjeligheden (figur 7-42). Samtidig falder protein- og vandindholdet i bladene – det betyder ringere fødekvalitet. Bladgnavende insekters fouragering skal derfor være synkroniseret med værtsplanternes sæsonudvikling.
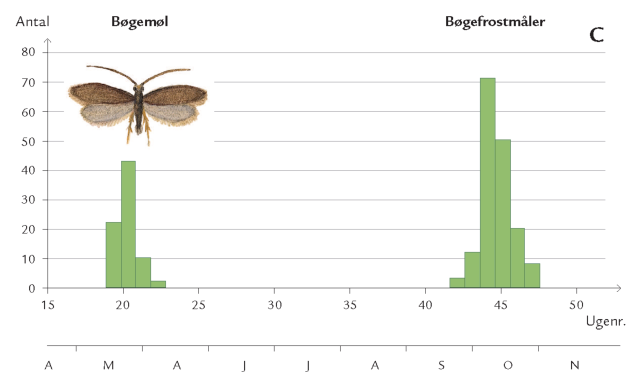
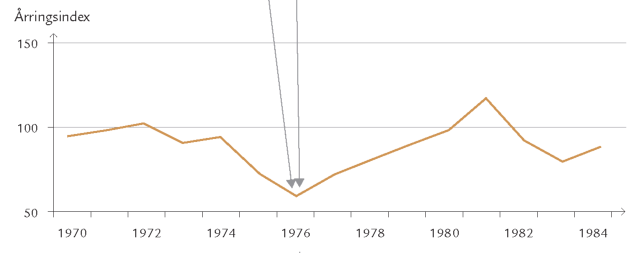
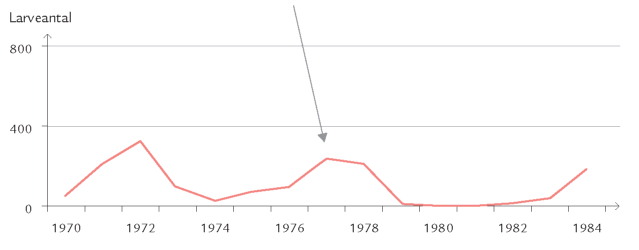
Et eksempel er frostmålerlarver, der begnaver blade af bøg, eg og andre løvtræer: Om efteråret klækker hunnerne fra skovbunden og kravler op ad stammerne for at lægge æg i barkrevner på grene og kviste (figur 7-35). Æggene klækker ved løvspring i april-maj. De nyklækkede larver skal hurtigst muligt trænge ind i en knop, der er ved at springe ud, for her kan de æde løs af de helt unge, delikate blade. Klækker æggene, før knopperne åbner sig, kan larverne ikke få adgang til bladene, men må forsøge at finde et andet træ med det rette knopstadium. De små larver spinder da en silketråd og lader sig drive med vinden (boks 7-5). Klækker æggene derimod et stykke tid efter, at bladknopperne er sprunget ud, kan bladene være for seje og fødekvaliteten for ringe på grund af et højt indhold af garvestoffer. Konsekvensen er da langsom larvevækst, øget dødelighed eller udvikling af unormalt små frostmålerhunner med lav ægproduktion. I Hestehaven var dødeligheden blandt æg og nyklækkede larver af frostmålere ca. 70 %.
Optimal larveudvikling er altså begrænset til et relativt snævert tidsinterval (figur 7-42). I udlandet har man studeret mekanismerne bag synkroniseringen mellem ægklækningstidspunkt og bladknoppernes udspring på individuelle egetræer. Man fandt bl.a., at frostmålerhunner, der er aktive tidligt om efteråret, oftest forekommer på egetræer, der springer tidligt ud.
Frostmålerhunner, som derimod er aktive senere på sæsonen, lægger fortrinsvis æg på egetræer, der springer sent ud. Da frostmålerhunnen er vingeløs, øges sandsynligheden for, at hun vil lægge æg på netop det træ, hvor hun tilbragte larvetiden, og som passer sammen med hendes nedarvede æglægningstidspunkt.
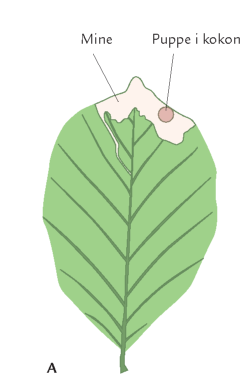
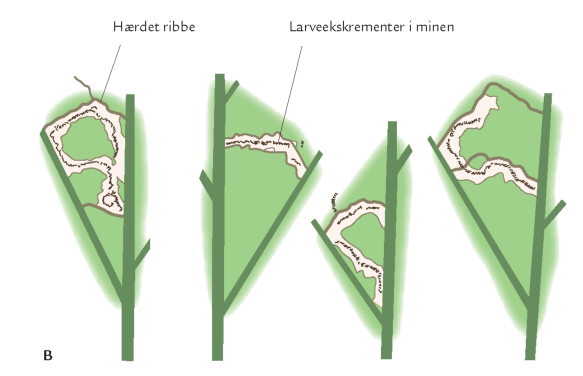
Også insektlarver, der minerer inde i bladkødet, er afhængig af god timing. Bøgeloppen (figur 7-44) lægger æg i bøgebladets midtribbe, og efter klækningen begynder larven at gnave en minegang inde i bladet (figur 7-43). Her æder bøgeloppelarven løs af bladkødet, producerer ekskrementer, vokser, skifter hud og forpupper sig. Omkommer den undervejs, forbliver den døde larve i minen. Bøgeloppeminer er derfor som en åben bog, hvor man kan kortlægge dødelighed og dødsårsager i en larvebestand. Det har bl.a. vist sig, at i visse år er bøgeloppens tidligste udvikling ikke godt nok synkroniseret med bøgebladets udvikling. I et køligt forår kan bøgeloppens æg- og larveudvikling forsinkes, samtidig hærder bladribberne, så de små larver ikke længere kan gennembryde dem; larverne dør af sult i den ufuldstændige mine (figur 7-43).
I et sådant år kan op til 70 % af de små larver omkomme af denne årsag.
Bladlus, småcikader og andre insekter med sugende munddele undgår at skulle håndtere de ufordøjelige plantekomponenter; de suger simpelthen plantesaft og celleindhold fra planten. Plantesaft er rig på sukkerstoffer, men fattig på kvælstof, så saftsugerne må indtage store mængder saft for at få dækket deres kvælstofbehov.
Nogle afgiver det overskydende, sukkerholdige vand som flydende ekstrementer – honningdug – en energikilde, der udnyttes af andre insekter, f.eks. sommerfugle, myrer og fluer.
Andre smådyr inducerer dannelsen af en abnorm vækst – en galle – på planten og skaffer sig på denne måde en næringsrig føderessource. Gallevæksten styres af en kemisk stimulus fra planteæderen (galmide, galmyglarve, galhvepslarve m.fl). De gallefremkaldende smådyr opholder sig inde i gallen, hvor de ernærer sig af det indre væv eller af plantesaften, der begge er letfordøjelige (figur 7-45).
Især stressede træer angribes
Skønt mange insekter æder af løvtræernes blade, er den samlede begnavning oftest lav.
I en 8-årig undersøgelse i Hestehaven blev der kun ædt 3-9 % af bøgens totale bladareal. En så lav begnavning er helt uden betydning for bøgetræernes fotosyntese, trivsel og produktion, men med års mellemrum kan både løvtræer og nåletræer udsættes for omfattende insektangreb og afløvning (se kapitlet Dynamik og forstyrrelser i skoven).
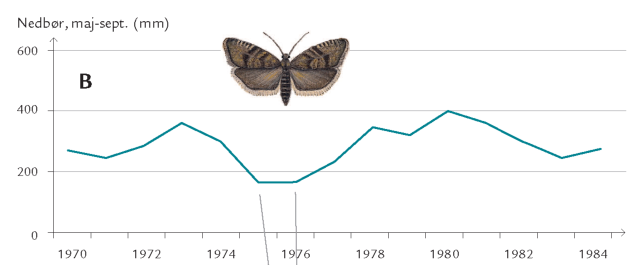
Hvad er baggrunden for disse udsving i planteæderbestande og angrebsniveau? Ofte er det et samspil mellem klimatiske faktorer, jordbundsforhold og værtstræets vitalitet. I almindelighed er træer, der ikke har optimale vækstforhold – stressede træer – mest udsat for insektangreb. Et eksempel er rød-gran og angreb af grannålevikleren (Epinotia tedella). De små sommerfugle lægger æg på grannåle, som larverne udhuler og spinder sammen i bundter (figur 7-46); om efteråret firer larverne sig ned til skovbunden, hvor de overvintrer. Viklerbestandens størrelse svinger, nu og da er der masseformering.
Mængden af sommernedbør menes at påvirke viklernes opformering. Det kan hænge sammen med, at sommertørke reducerer rød-granernes vækst, modstandsdygtighed og produktion af forsvarsstoffer mod insektangreb. Det giver viklerlarverne bedre levevilkår på de stressede træer og flere vil overleve til det følgende forår, hvor de mange nyklækkede sommerfugle vil lægge mange æg på grannålene. Larverne vil stortrives på de svækkede træer, og larvebestanden vil nå nye højder. Effekten på larvebestanden af svigtende sommernedbør viser sig således med et års forsinkelse.









































Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.