I de fleste jordbunde har det vist sig, at dyrene, der samlet kaldes faunaen, kun står for få procent af jordbundens samlede energiomsætning.
I Hestehaven (boks 7-1 i Liv og samspil i jordbunden) blev det således beregnet, at faunaen kun var ansvarlig for at nedbryde en stofmængde, der svarer til ca. 3 % af den energi, der hvert år blev tilført jordbunden med døde plantedele. Resten sørgede svampe og bakterier for.
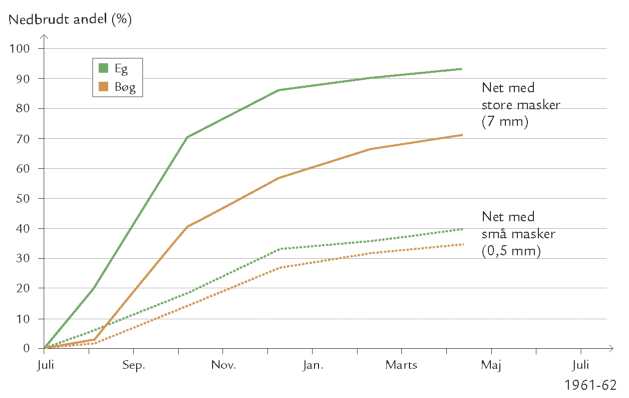
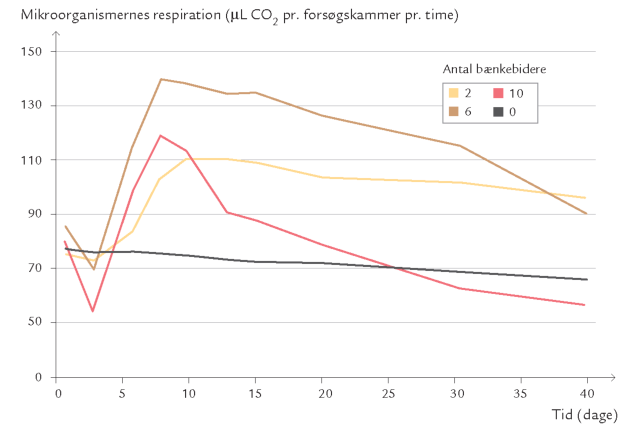
Da kun få dyr kan betragtes som primære nedbrydere og ofte lever af at æde svampe og bakterier – dvs. som sekundære nedbrydere skulle man umiddelbart tro, at de havde ringe eller eventuelt negativ indvirkning på nedbrydningsprocessen. Talrige forsøg i laboratoriet og i naturen har imidlertid vist, at det modsatte er tilfældet, dvs. at nedbrydningen af f.eks. døde blade går langsommere, hvis der ikke er svampeeller bakterieædende dyr til stede (figur 10-10 og 10-11).
Dyrenes aktivitet synes således at stimulere nedbrydningsprocessen. I de følgende afsnit uddybes de forskellige måder, stimuleringen foregår på. Der er dog også tilfælde, hvor et for stort antal dyr kan hæmme nedbrydningen, hvilket kan skyldes, at primærnedbrydernes vækst ikke kan følge med dyrenes fortæring.
Mekaniske virkninger på udgangsmaterialet og spredning af sporer og kim
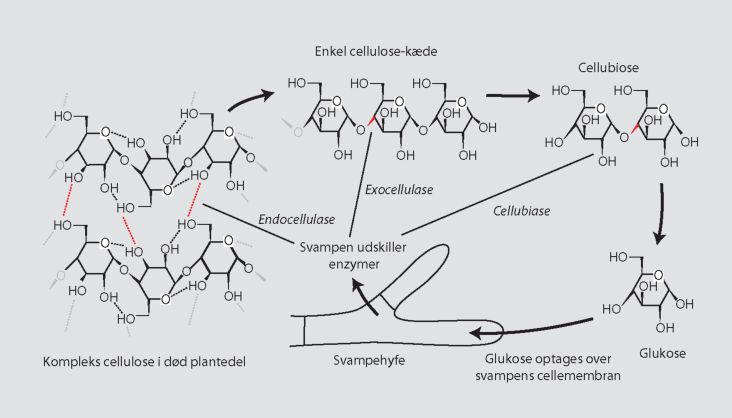
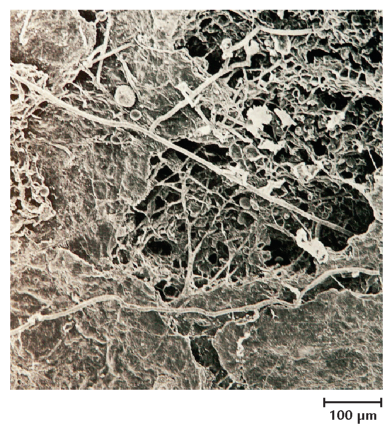
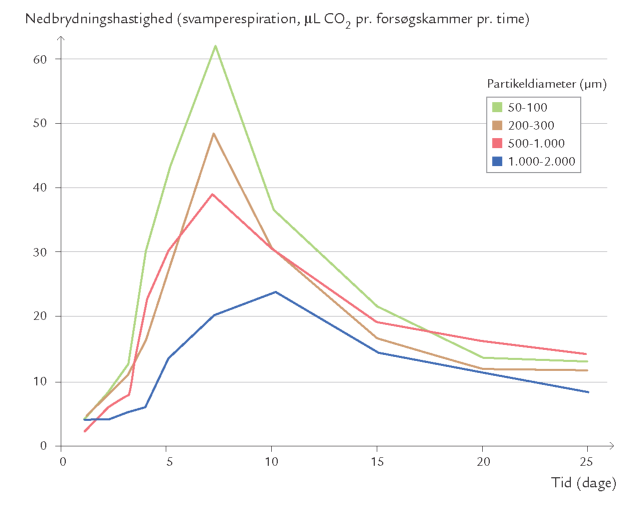
Rent mekanisk betyder dyrenes ædeaktivitet, at planterester og andet organisk materiale findeles. Derved blottes nye og større overflader, som kan udnyttes af svampe og bakterier (figur 10-11). I laboratorieforsøg med broget læderporesvamp podet på findelte egeblade har det vist sig, at nedbrydningsaktiviteten stiger med øget findeling af bladene (figur 10-12). Jo større overflade, desto større nedbrydningsaktivitet. I dødt træ fremmer tunnelborende dyr primærnedbrydernes adgang til det døde ved.
Det er også af stor betydning, at dyrene på deres overflade og i tarmen transporterer sporer og kim af svampe og bakterier, som på den måde spredes til steder i jorden og i døde plantedele, som de ellers vanskeligt kan kolonisere (figur 10-13). F.eks. vandrer mange jordbundsdyr op og ned i jorden, bl.a. som følge af svingende fugtighedsforhold, og de kan transportere kim af svampe og bakterier fra et lag af jorden til et andet. Sådanne transporter har f.eks. betydning for koloniseringen af nyfaldet løv med svampe og bakterier fra dybere jordlag.
Mikrogræsningens virkning på svampe- og bakterievækst
Jordbundsdyrenes fortæring af svampehyfer og bakteriekolonier kan som nævnt stimulere disse organismers vækst og stofskifte. Det skyldes bl.a., at svampehyfers og bakteriekoloniers vækst kan gå i stå, fordi de har optaget alle de vigtige næringsstoffer, f.eks. kvælstof, i substratet. Når så dyrene æder dem, bliver hovedparten af disse stoffer udskilt igen, så de kan udnyttes af nye generationer af bakterier og svampe – eller af planternes rødder (figur 10-14).
Substratet, dvs. det døde organiske materiale med indhold af organismer, bliver faktisk mere og mere rigt på næringsstoffer under nedbrydningen, fordi kulstoffet forbrændes af organismerne og forsvinder i kuldioxid, mens næringsstofmolekylerne, f.eks. kvælstof, stort set bevares i substratet. Forholdet mellem kulstof og kvælstof (C:N-forholdet) i et bøgeblad falder således under nedbrydningen fra ca. 38 i de nyfaldne blade til ca. 10, hvilket svarer til C:N-forholdet i svampe og bakterier. Først da bliver kvælstof ikke længere begrænsende for disse organismernes vækst, og der bliver et overskud af simple kvælstofforbindelser, som ikke straks igen bliver bundet i levende celler. De overskydende kvælstofforbindelser kan optages af planterødder eller eventuelt udvaskes til grundvandet.
En mere direkte form for vækststimulering er fundet i samspillet mellem den svampeædende porespringhale Protaphorura armata og koblingssvampen Mortierella isabellina. Det, at springhalen åd af svampehyferne, førte til, at de udviklede hyfeafsnit, som voksede særlig hurtigt, og som producerede nedbrydningsenzymer (protease- og alfa-amylase) i langt større udstrækning end de “normale” hyfer. Den øgede hyfevækst kom dyrene og nedbrydningen til gode, men det er tvivlsomt, om der samlet set var en positiv virkning for svampen, for dens sporeproduktion blev mindre.
Svampe og bakterier i jordbundsdyrenes tarm og ekskrementer
En vigtig pointe i forbindelse med dyrenes betydning for nedbrydningsprocessen er som omtalt ovenfor, at mange bakterier og svampesporer kan overleve i dyrenes tarm (figur 10-13). En stor del af svampesporerne er spiringsdygtige, når tarmindholdet afgives som ekskrementer, og de næringsrige ekskrementer er et godt medium for fremvækst af ny svampehyfer. Bakterierne kan, som det uddybes nedenfor, endog opformeres under tarmpassagen, og både bakterier og mikrosvampe udvikler sig ofte bedre i dyrenes ekskrementer end i den omgivende jord.
Bortset fra nogle få vedædende insekter har nedbryderfaunaen ikke en egentlig, specialiseret tarmflora. I stedet sker der en opformering af jordbundsbakterier i tarmen. Hos regnorme, men også hos f.eks. tusindben, der æder dødt organisk materiale, har man fundet, at bakteriernes antal øges under opholdet i tarmen. Hos kugletusindbenet Glomeris marginatahar forsøg vist, at antallet af bakterier voksede fra 400 millioner pr. gram af den indtagne føde til 2.300 millioner pr. gram af tarmindholdet og endelig til 40.000 millioner pr. gram ekskrementer. Bestandstætheden af bakterier var altså vokset 100 gange ved passage gennem dyret. Årsagen hertil kan være, at tarmindholdet er mere næringsrigt end det gennemsnitlige organiske materiale i jorden, da dyrene vælger at æde det bedste, der umiddelbart er til rådighed, og at det beskyttede miljø i tarmen er væsentligt mere gunstigt for bakterierne end i den omgivende jordbund. Virkningen er ikke den samme for alle bakterier, og tarmpassagen kan især gavne bakterier, som forekommer i lille antal i dyrenes omgivelser. Resultatet er, at nogle arter af bakterier i føden udnyttes som næringskilde for dyrene, mens andre trives i tarmen og opformeres stærkt.
Svampehyfer er i øvrigt også en vigtig bestanddel af tusindbenenes føde, men her falder antallet af hyfestykker markant ved passage gennem dyret. Der bliver derfor en anden mængde og sammensætning af bakterier og svampe i ekskrementerne end i den indtagne føde.
På grund af den berigelse med mikroorganismer og let fordøjeligt stof, der finder sted under og efter tarmpassagen, er ekskrementer ofte en bedre fødekilde end den del af jorden, som ikke har været ædt. Konsumering af egne eller andre dyrs ekskrementer (koprofagi) er derfor meget udbredt blandt jordbundens dyr, f.eks. regnorme, bænkebidere, tusindben, springhaler og pansermider. Det er ofte mindre arter, som æder større arters ekskrementer, hvorved den samme mængde jord bliver ædt gang på gang. Den gentagne koprofagi, hvor det samme materiale gang på gang ædes, virker som en forlængelse af fordøjelsesprocessen i tarmen, hvilket resulterer i en effektiv nedbrydning af det døde plantemateriale.
Skønt ekskrementer af jordbundsdyr generelt viser stor biologisk aktivitet og derfor nedbrydes hurtigere end det organiske materiale, som ikke har været gennem dyrenes tarm, er der undtagelser, hvor nedbrydningshastigheden nedsættes og det organiske stof stabiliseres i ekskrementerne. Det gælder f.eks. hos regnorme, hvor der i tarmen dannes stabile forbindelser mellem lermineraler og organisk stof, hvilket resulterer i dannelsen af jordkrummer, som har stor betydning for jordens evne til at binde næringsstoffer og vand – men som altså ikke nedbrydes yderligere.
Man har også konstateret, at ekskrementer af springhaler og mider kan være meget svært nedbrydelige. Blandt miderne kan nævnes visse pansermider, der lægger æg på grannåle, hvorefter ungerne udhuler nålene. Mod forventning taber de nåle, der udhules af miderne, vægt langsommere end de ikke angrebne nåle, i hvert fald i de første 3 år af nålenes meget langvarige nedbrydning. Det skyldes, at midernes ekskrementer er mere resistente mod nedbrydning end vævet i grannålene, og sådanne ekskrementer er formodentlig med til at danne jordbundens stabile humusforbindelser.
Regnormenes betydning for nedbrydningen
Regnormenes rolle i nedbrydningen er knyttet til deres stimulering af de primære nedbrydere, svampe og bakterier.
I muldrige løvskove kan regnorme fjerne en stor del af den årlige produktion af nedfaldsløv. En bestand af stor regnorm i en blandingsskov kan i løbet af ca. 3 måneder konsumere et årligt løvfald på 3 tons pr. ha. Regnorme producerer derfor generelt store mængder ekskrementer. På gode, muldrige lokaliteter med bestande på 200-400 individer pr. m2 er der målt en årlig produktion af ekskrementer, der varierer fra 1-26 kg pr. m2eller op til ca. 260 tons pr. ha.
Ekskrementerne indeholder en forholdsmæssigt større mængde næringsstoffer som kulstof, kvælstof og fosfor end den omgivende jord. Det skyldes som også nævnt tidligere, at regnorme aktivt opsøger føde med højt indhold af organisk materiale.
Det er især de store, dybtgravende (aneciske) regnormearter, der producerer betydelige mængder ekskrementer på selve jordoverfladen (figur 10-15, se også boks 7-2 i Liv og samspil i jordbunden). De mindre overfladelevende (epigæiske) arter danner ekskrementer, der hovedsageligt består af fragmenteret løvmateriale, som inkorporeres i førnen. Endelig afsætter de såkaldt jordlevende (endogæiske) arter, der normalt konsumerer betydelige mængder mineraljord sammen med fødematerialet, deres ekskrementer i gangsystemer og hulrum nede i jorden.
Regnormeekskrementer består i varierende grad af mineraljord og rester af findelt og fordøjet plantemateriale, der er bundet sammen i mere eller mindre stabile jordkrummer (1-10 mm). Disse jordkrummer har en betydelig vandstabilitet, der bl.a. opnås ved cementering og sammenbinding af partiklerne vha. slim fra regnormene og bakterier, plantefibre og svampehyfer samt kemiske bindinger mellem organisk stof og mineraler. Regnormeekskrementer bidrager hermed i udtalt grad til opretholdelse af jordens system af porer og dermed dens vandbindingsevne. En sådan optimal jordstruktur, som sikrer passende fugtighed, udluftning og mulighed for passage af planterødder, svampehyfer og dyr, er af stor betydning for en effektiv nedbrydning og recirkulering af næringsstoffer i jordbunden.
Regnormenes betydning for dannelsen af hhv. muld og mor er omtalt i Jordbundens lag













Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.